
Susceptibility status of major malaria vectors to novaluron, an insect growth regulator South-Eastern Tanzania
Amos Justinian Ngonzi, Letus Laurian Muyaga, Halfan Ngowo, Naomi Urio, John-Mary Vianney, Dickson Wilson Lwetoijera
Corresponding author: Amos Justinian Ngonzi, Ifakara Health Institute, Environmental Health and Ecological Science Department, PO Box 53, Ifakara, Tanzania 
Received: 15 Feb 2022 - Accepted: 21 Mar 2022 - Published: 05 Apr 2022
Domain: Public health
Keywords: Insect growth regulator, novaluron, susceptibility, An. arabiensis, An. gambiae, An. funestus, Tanzania
©Amos Justinian Ngonzi et al. Pan African Medical Journal (ISSN: 1937-8688). This is an Open Access article distributed under the terms of the Creative Commons Attribution International 4.0 License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Cite this article: Amos Justinian Ngonzi et al. Susceptibility status of major malaria vectors to novaluron, an insect growth regulator South-Eastern Tanzania. Pan African Medical Journal. 2022;41:273. [doi: 10.11604/pamj.2022.41.273.33793]
Available online at: https://www.panafrican-med-journal.com//content/article/41/273/full
Research 

Susceptibility status of major malaria vectors to novaluron, an insect growth regulator South-Eastern Tanzania
Susceptibility status of major malaria vectors to novaluron, an insect growth regulator South-Eastern Tanzania
Amos Justinian Ngonzi1,2,&, Letus Laurian Muyaga1, ![]() Halfan Ngowo1,
Halfan Ngowo1, ![]() Naomi Urio1,2,
Naomi Urio1,2, ![]() John-Mary Vianney2, Dickson Wilson Lwetoijera1,2
John-Mary Vianney2, Dickson Wilson Lwetoijera1,2
&Corresponding author
Introduction: application of Insect Growth Regulator (IGR) such as pyriproxyfen has shown a promising result in controlling malaria transmitting mosquitoes through autodissemination technique. Novaluron that inhibits the chitin development at mosquito larval stage present a promising candidate IGR for rotation with pyriproxyfen to prevent a chance of resistance development. This study assessed the susceptibility of immature stages of Anopheles arabiensis, Anopheles gambiae and Anopheles funestus to novaluron.
Methods: susceptibility bioassays using technical grade novaluron (98% active ingredient) were performed inside the semi-field system using first instar larvae of Anopheles species. For each tested species, a total of 1500 larvae were used in the bioassay. Concentration range of 0.01 mg/l to 2 mg/l of novaluron were tested to establish Lethal Concentration (LC) sufficient to kills 50%, 90% and 99% of the exposed larvae by using log-dose response analysis.
Results: of the tested mosquitoes, Anopheles gambiae were highly susceptible to novaluron followed by An. arabiensis and then An. funestus. Lethal concentrations, LC50, LC90 and LC99 (95%CI) in mg/l for An. gambiae were 0.018, 0.332 and 2.001 respectively. For An. arabiensis were 0.026, 0.546 and 2.013; and for An. funestus were 0.032, 1.00 and 5.580. High larval mortality was recorded at high concentration (2mg/L), with 80% mortality within 3 days post exposure.
Conclusion: the study demonstrates the efficacy of novaluron in controlling Anopheles mosquito species at immature stages via larval mortality. These findings warrant further testing of novaluron for autodissemination by different vector species for its inclusion in rotation to prevent development of resistance.

Outdoor and indoor malaria transmissions have profoundly led to the present malaria morbidity and mortality. In the year 2020 alone, there was 241 million malaria cases and 627,000 deaths globally [1]. Disproportionately, countries in sub-Saharan Africa, including Tanzania, have continue to accounting 95% of total cases and 602,000 deaths [1]. Additionally, malaria is considered to be a major economic burden in Africa, whereby the continent lost 12 billion USD in year 2000 [2]. It was demonstrated that, 10% global decrease in malaria incidence can result up to 0.3% average increase in income per capita, with high malaria endemic areas benefiting most [3,4]. With this economic impact, malaria prevention strategies are highly needed for implementation to overcome the burden.
Different strategies for malaria prevention with vector control tools, primarily the long-lasting insecticide mosquito nets (LLINs) and indoor residual sprays (IRS) are strongly recommended by the Word Health Organization [1]. Across sub-Saharan Africa where >90% of the disease burden is concentrated, both LLINs and IRS have significantly suppressed malaria vectors, especially, those that bite and rest indoors. These preventions and controls measures have contributed nearly 40% of 57% reduction of clinical disease incidences [5]. However, rapid increase in insecticide resistance and observed higher outdoor biting and resting patterns of malaria vectors jeopardize future application of these interventions towards malaria elimination efforts [6]. These challenges demonstrate urgent need for alternative malaria vector control measures which can complement the existing malaria vector interventions.
Larval source management, is another vector control tool that uses chemical and biological agents used to control malaria vectors in aquatic stages [7,8]. This technique works by reducing vector densities at mosquito breeding habitats either through killing effect of mosquito immature stages or adult emergence inhibition effect [7,8]. Despite of the success attained by larviciding for controlling malaria vectors across Africa, high operational cost and low coverage of the targeted breeding habitats remain to be the greatest challenges [9-12].
Of importance, mosquito assisted larviciding commonly known as autodissemination with insect growth regulators (IGRs) (i.e pyriproxyfen and novaluron) can accurately target aquatic habitats with larvicide and deliver the desired impact [13-16]. By definition, auto-dissemination is the management method in which insects such as mosquitoes get exposed and pick biological or chemical insecticide such as IGRs while seeking the host or feeding or resting, and transfer lethal concentrations vertically or horizontally to the oviposition sites and result in reduction of adult mosquitoes [13,17].
Novaluron is an IGR that have been recently tested using autodissemination technique against different mosquito species [16]. It inhibits the chitin synthesis process at larval stages of mosquitoes through contact and ingestion of a benzoylphenyl urea formation, whereby larvae succumb to death as the results of abnormal endocuticle deposition [16,18-20]. Of interest, novaluron has a reduced risk to the environment including mammals, birds, aquatic animals and non-targeted insects [19,20].
Recently, novaluron has shown effect in reducing adult mosquito density at their larval stage. With evidences reported on its efficacy, novaluron has excellently worked against immature Anopheles quadrimaculatus, Aedes aegypti and Culex quinquefasciatus in the laboratory and field settings [16,21,22]. Likewise, using autodissemination technique, Swale et al. 2018 demonstrated the effect of novaluron against An. quadrimaculatus, causing up to 22% reduction in adult emergence as the results of larval mortality [16]. Despite the benefits that novaluron offers to control other mosquito borne diseases, there is no evidence of its application to control the main malaria vectors in rural South-eastern Tanzania. Here, we evaluated the susceptibility of the main malaria vectors in South-eastern Tanzania, Anopheles arabiensis, Anopheles gambiae and Anopheles funestus, to varying doses of novaluron under semi-field setting.

Study site: this study was conducted in a Semi-Field System (SFS) of Ifakara Health Institute between July-September, 2021. The experimental SFS is located at Kining´ina village (8.11417°S, 36.67484°E) in Ifakara, Kilombero District, South-eastern Tanzania (Figure 1). Detailed description and dimensions of the SFS has been described elsewhere [14,23].
Mosquitoes: the study used insectary reared mosquitoes from the established colonies of An. arabiensis, An. gambiae and An. funestus. Details of colonies´ rearing and maintenance procedures are provided elsewhere [23-26]. All bioassays used first instar larvae owing to its high susceptibility to novaluron [16].
Preparation of test concentrations: test concentrations of novaluron, 98% test concentration (technical materials; Jiaozuo Huisell Chem, Ltd, China) were prepared using standardized procedures [27]. Mass of novaluron; 0.01mg, 0.05mg, 0.1mg and 2.0 mg were measured using electronic beam balance and dissolved in 1000 ml of tap water to prepare the concentrations; 0.01mg/L, 0.05mg/L, 0.1mg/L and 2mg/L respectively. Aliquots of 200 ml of each prepared concentration was placed in plastic cup (four replicates) for bioassays plus four control plastic cups containing tap water alone.
Laboratory susceptibility test: the bioassays had control and treatment cups containing test concentration and mosquito larvae. The expected outcome was larval mortality at the treatment cups compared to the control cups to confirm lethal concentrations that is required to kill 50%, 90% and 99% of exposed larvae. Twenty-five (25) first instar larvae per replicate were exposed to novaluron concentrations; 0.01mg/L, 0.05mg/L, 0.1mg/L and 2.0 mg/L. The set-up was repeated three times on different days to counter confounders in the bioassay. Larvae were fed at 1-day interval with Tetramin� fish food throughout the course of the assay. The larval mortality was monitored on 24 hours interval until all larvae were dead or pupated. Dead larvae were counted and removed from the plastic cups. The larval mortality data was corrected using Abbott´s formula. Log-dose response analysis was carried out to determine lethal concentration of 50%, 90% and 99% (LC50, LC90 and LC99). The temperature during the assay ranged between 24-27°C, 80% ± 10% relative humidity and the photoperiod of 12L: 12D. Diagnostic concentration was established from the lethal concentrations that killed up 99% of the exposed Anopheles larvae, and it was defined as the two times of LC99 [27].
Effect of novaluron on pupation rate: the effect of novaluron on larval mortality was recorded to determine the percentage inhibition of pupation (PI%). Moribund and dead larvae and pupae that did not completely separated from the larvae case, were considered as affected by novaluron. The experiment ended 15 days post-exposure. The data from all replicates were combined to calculate the mean of affected larvae. The PI% of Anopheles larvae caused by novaluron was calculated using the formula:
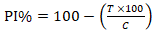
Whereby; T= percentage pupation in treated cups; C= percentage pupation in control cups.
Statistical analysis: data were analyzed using R software (Rv-4.1.1) [28]. Generalized linear mixed models were used to assess the proportion of dead larvae for each concentration [29]. The proportion of dead larvae were modelled as a response variable and test concentrations were considered as fixed effect while replicates and days were included as a random term to account for the pseudo replicates and unexplained variation between days. Lethal concentrations, LC50, LC90 and LC99 were determined using log-dose response analysis from dose-response curve package [30]. The curve was used to determine the desired concentration of novaluron. Additionally, Tukey honest significance test (TukeyHSD) was used to assess the pairwise difference between different concentration levels. Risk ratio and their corresponding 95% CI were reported, whereby, the statistical significance was considered when p-values ≤ 0.05.
Ethical consideration: prior to laboratory work the research proposal was presented to the Nelson Mandela Institute of Science and Technology and Ifakara Health Institute for approval. Further, the ethical approval for the study was granted by Institutional Review Board of Ifakara Health Institute (IHI/IRB/No: 20-2021).
Laboratory susceptibility test: high larval mortality was recorded with high concentrations of novaluron, whereas low concentrations were associated with delayed mortality. An. gambiae larvae were more susceptible with LC50 and LC90 being 0.0179 mg/L and 0.332 mg/L respectively, while LC50 and LC90 for An. arabiensis and An. funestus was 0.02561 mg/L and 0.5460 mg/L; and 0.0323 mg/L and 1.000mg/L respectively (Table 1). Larval mortality in the respective control ranged from 7% to 15% depending on mosquito larvae species exposed (Figure 2). However, the laboratory susceptibility test yielded the diagnostic concentrations for all three-target species (Table 1). The results on the comparison of different concentrations on the larval mortality are summarized in Table 2. In three Anopheles species, An. gambiae was highly susceptible [RR = 1.0842, p < 0.005], followed by An. arabiensis [RR = 10.237, p < 0.001], and An. funestus [RR = 11.41, p < 0.001] at test concentration of 0.1 mg/L (Table 2). However, the pair-wise comparison test using Tukey´s HSD showed significant difference between control and 0.1 mg/L for An. arabiensis (z = 42.83, p < 0.001), An. gambiae (z = 43.87, p < 0.001) and An. funestus (z = 40.53, p < 0.001) (Figure 3).
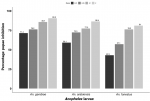
Effect of novaluron on pupation rate: the results demonstrated high percentage inhibition of pupation with increase in concentration. Highest PI% was recorded at 2 mg/L compared to other low concentrations across all three Anopheles species with PI% of 90.5%, 86.4% and 81.0% for An. gambiae, An. arabiensis and An. funestus respectively (Figure 4).
This study demonstrated up to 80% and 90% larval mortality and pupae inhibition of the exposed larvae of Anopheles gambiae, Anopheles arabiensis and Anopheles funestus, an afro-tropical malaria vectors to novaluron under controlled settings. These findings corroborate other previous reports that demonstrated the control of Anopheline, Adenine and Culicine mosquitoes using novaluron under laboratory and field settings [16,18,31]. Lethal concentrations sufficient to kills 50% and 90% of the exposed mosquito larvae were different across three tested species; all achieved within 15 days post-exposure. This highlights delayed developmental duration of exposed larvae as the results of novaluron effect [18,32,33]. Over 50% mortality of all Anopheles larvae were observed in between 2nd and 3rd day post-exposure at maximum test concentration of novaluron (2mg/L). Despite of the development of the exposed larvae to 3rd instar, none was able to reach 4th instar or pupae stage. Previous studies assessing the effect of novaluron to mosquito larvae have also reported slow and extended larval growth and delayed mortality post-exposure time [16,18,21]. This delayed mortality is expected to reduce pressures on mosquitoes to develop resistance to the novaluron, and offer a more sustainable insecticide for vector control thereof [21,32].
In comparison, An. gambiae was more susceptible to novaluron followed by An. arabiensis and lastly An. funestus. Lethal concentrations of novaluron required to kill 50%, 90% and 99% of An. funestus larvae was one to two and half times higher than that for An. gambiae and An. arabiensis. In addition, the diagnostic concentration for An. funestus (11.160 mg/L) was three times higher than that of An. gambiae (4.002 mg/L) and An. arabiensis (4.026 mg/L). Although not investigated under this study, the probable cause for reduced susceptibility might be a high level of pyrethroids resistance in An. funestus documented by other studies in the same study location [34,35]. A single study, has also highlighted possibility of cross-resistance between pyrethroids and insect-growth regulators within Anopheles population, which might be applicable in this case [36]. The difference of lethal and diagnostics concentrations recorded under different studies might be explained by physiological difference with test species [16,18].
There is increasing evidence that the use of IGRs of different mode of action against mosquitoes can counteract and/or delay the development of insecticide resistance [8,37]. These findings point out the efficacy of novaluron in reducing adult mosquito population at breeding habitats. Thus, an additional insecticide that may be applied in rotation with other IGRs, such as pyriproxyfen to manage insecticide resistance and reduce adult mosquito population at their larval habitats. Of importance, World Health Organization (WHO) approval on the use of novaluron in drinking water signals its safety to human and animals, and warrant its testing using conventional larviciding or autodissemination techniques in different settings [38].
This study had a number of limitations; under laboratory settings no attempt was made to test for persistence of novaluron in the test cups beyond single larval exposure. While low susceptibility of An. funestus to novaluron was attributed to its high insecticide resistance status, no actual experiments that were carried to ascertain this assertion, and this represent another study limitation. Therefore, these limitations add on the list of future studies towards development of novaluron as the potential larvicide for malaria vector control.
This study conclude that major malaria vectors found in Kilombero, Tanzania are susceptible to novaluron at low concentration. This is the first demonstration on the susceptibility of An. gambiae, An. arabiensis, An. funestus to novaluron under laboratory settings. These results warrant further semi-and field testing of novaluron using the autodissemination technique against An. gambiae, An. arabiensis and An. funestus for its inclusion in rotation to prevent evolution of insecticide resistance.
What is known about this topic
- Susceptibility of other mosquito species of disease importance to novaluron;
- Possibility of autodissemination of novaluron by other mosquitoes such as Anopheles quadrimaculatus.
What this study adds
- Demonstrated that major malaria vectors in Tanzania are highly susceptible to low dosages of novaluron;
- Offer empirical evidence on novaluron as an additional IGR for malaria vector control at its aquatic habitats, that can be considered for application in rotation with other IGR such as pyriproxyfen (PPF) to manage insecticide resistance development.
The authors declare no competing interests.
Amos Justinian Ngonzi was involved in conceptualization and designing of the study, conducting experiment, data collection, data management and analysis, and writing the manuscript. Dickson Wilson Lwetoijera and John-Mary Vianney were involved in conceptualization study and its design, data analysis, supervision and manuscript revision. Halfan Ngowo, Letus Laurian Muyaga and Naomi Urio were involved in study design, data analysis and manuscript write-up. All authors read and approved the final manuscript.
On their consent, we thank Ms. Monica Mpingwa, Ms. Zuhura Kombo and Ms. Nuru Nchimbi for the support of maintaining mosquito colonies. We also thank Mr. Betwel John Msugupakulya and Ms. Jacqueline Mgaya for the supportive guidance in setting up the bioassays.
Table 1: larval susceptibility of malaria vector species to novaluron
Table 2: larvae mortality and their risk effects at different concentrations of novaluron
Figure 1: A) semi-field system used in experiments; B) chambers inside the semi-field; C) mosquito rearing insectary inside semi-field system
Figure 2: cumulative mortality percentage of: A) An. gambiae; B) An. arabiensis and C) An. funestus larvae when 1st instar larvae were treated with novaluron-chitin synthetic inhibitor
Figure 3: results of pair-wise post hoc comparison using Tukey's honestly significance tests (Tukey's HSD); similarities and differences between larvae mortality at different concentrations; A) An. gambiae; B) An. arabiensis and C) An. funestus
Figure 4: percentage inhibition of pupation of different malaria vectors at different test concentration of novaluron 15 days post-treatment
- World Health Organization. World malaria report. 2021. Accessed October 29, 2020.
- World Health Organization. The Africa malaria report. 2003. Accessed October 29, 2020.
- Gallup JL, Sachs JD. The economic burden of malaria. Am J Trop Med Hyg. Jan-Feb 2001;64(1-2 Suppl):85-96. PubMed | Google Scholar
- Sarma N, Patouillard E, Cibulskis RE, Arcand JL. The economic burden of malaria: revisiting the evidence. Am J Trop Med Hyg. 2019;101(6):1405-1415. PubMed | Google Scholar
- Bhatt S, Weiss DJ, Cameron E, Bisanzio D, Mappin B, Dalrymple U et al. The effect of malaria control on plasmodium falciparum in Africa between 2000 and 201 Nature. 2015;526(7572):207-211. PubMed | Google Scholar
- Hemingway J. The way forward for vector control. Science. 2017;358(6366):998-999. PubMed | Google Scholar
- Walker K, Lynch M. Contributions of anopheles larval control to malaria suppression in tropical Africa: review of achievements and potential. Medical and Veterinary Entomology. 2007;21(1):2-21. PubMed | Google Scholar
- Tusting LS. Larval source management: a supplementary measure for malaria control. Outlooks Pest Manag. 2014;25(1):41-43. Google Scholar
- Chaki PP, Govella NJ, Shoo B, Hemed A, Tanner M, Fillinger U et al. Achieving high coverage of larval-stage mosquito surveillance: challenges for a community-based mosquito control programme in urban Dar es Salaam, Tanzania. Malar J. 2009 Dec 30;8:311. PubMed | Google Scholar
- IVCC. Vector control 10 challenges and 10 solutions. 2020. Accessed October 29, 2020.
- Stanton MC, Kalonde P, Zembere K, Hoek Spaans R, Jones CM. The application of drones for mosquito larval habitat identification in rural environments: a practical approach for malaria control? Malar J. 2021;20(244). Google Scholar
- Geissbühler Y, Kannady K, Chaki PP, Emidi B, Govella NJ, Mayagaya V et al. Microbial larvicide application by a large-scale, community-based program reduces malaria infection prevalence in urban Dar Es Salaam, Tanzania. PLoS One. 2009;4(3):e5107. PubMed | Google Scholar
- Devine GJ, Perea EZ, Killeen GF, Stancil JD, Clark SJ, Morrison AC. Using adult mosquitoes to transfer insecticides to Aedes aegypti larval habitats. Proc Natl Acad Sci U S A. 2009;106(28):11530-11534. PubMed | Google Scholar
- Lwetoijera D, Harris C, Kiware S, Dongus S, Devine GJ, McCall PJ et al. Effective autodissemination of pyriproxyfen to breeding sites by the exophilic malaria vector Anopheles arabiensis in semi-field settings in Tanzania. Malar J. 2014 Apr 29;13:161. PubMed | Google Scholar
- Mbare O, Lindsay SW, Fillinger U. Testing a pyriproxyfen auto-dissemination station attractive to gravid Anopheles gambiae sensu stricto for the development of a novel attract-release -and-kill strategy for malaria vector control. BMC Infect Dis. 2019 Sep 11;19(1):800. PubMed | Google Scholar
- Swale DR, Li Z, Kraft JZ, Healy K, Liu M, David CM et al. Development of an autodissemination strategy for the deployment of novel control agents targeting the common malaria mosquito, anopheles quadrimaculatus say (Diptera: Culicidae). PLoS Negl Trop Dis. 2018 Apr 11;12(4):e0006259. PubMed | Google Scholar
- Gaugler R, Suman D, Wang Y. An autodissemination station for the transfer of an insect growth regulator to mosquito oviposition sites. Med Vet Entomol. 2012;26(1):37-45. PubMed | Google Scholar
- Arredondo-Jiménez JI, Valdez-Delgado KM. Effect of Novaluron (Rimon� 10 EC) on the mosquitoes Anopheles albimanus, Anopheles pseudopunctipennis, Aedes aegypti, Aedes albopictus and Culex quinquefasciatus from Chiapas, Mexico. Med Vet Entomol. 2006;20(4):377-387. PubMed | Google Scholar
- Dhadialla TS, Retnakaran A, Smagghe G. Insect growth- and development-disrupting insecticides. In: Gilbert LI, editor. Insect development: morphogenesis, molting and metamorphosis. London, UK: Academic Press; 2009;679-740. Accessed October 29, 2020.
- Tunaz H, Nedim U. Insect growth regulators for insect pest control. 2004. Accessed October 29, 2020.
- Mulla MS, Thavara U,Tawatsin A, Chompoosri J, Zaim M, Su T. Laboratory and Field Evaluation of Novaluron a New Acylurea Insect Growth Regulator against Aedes aegypti ( Diptera: Culicidae ). J Vector Ecol. 2003 Dec;28(2):241-54. PubMed | Google Scholar
- Su T, Cheng M, Melgoza A, Thieme J. Laboratory and field evaluations of mosquiron�. 12CRD, a new formulation of Novaluron, against Culex mosquitoes. 2014;30(4):284-290. PubMed | Google Scholar
- Ferguson HM, Ng'habi KR, Walder T, Kadungula D, Moore SJ, Lyimo I et al. Establishment of a large semi-field system for experimental study of African malaria vector ecology and control in Tanzania. Malar J. 2008 Aug 20;7:158. PubMed | Google Scholar
- Agumba S, Gimnig JE, Ogonda L, Ombok M, Kosgei J, Munga S et al. Diagnostic dose determination and efficacy of chlorfenapyr and clothianidin insecticides against Anopheles malaria vector populations of western Kenya. Malar J. 2019 Jul 17;18(1):243. PubMed | Google Scholar
- Ngowo HS, Hape EE, Matthiopoulos J, Ferguson HM, Okumu FO. Fitness characteristics of the malaria vector Anopheles funestus during an attempted laboratory colonization. Malar J. 2021 Mar 12;20(1):148. PubMed | Google Scholar
- Ng'habi KR, Lee Y, Knols BGJ, Mwasheshi D, Lanzaro GC, Ferguson HM. Colonization of Malaria vectors under semi-field conditions as a strategy for maintaining genetic and phenotypic similarity with wild populations. Malar J. 2015;14(10):1-11. Google Scholar
- WHOPES. Guidelines for laboratory and field testing of mosquito larvicides. World Health Organization. 2005;1-41. Google Scholar
- Team RC. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2021;3.
- Bates D, Machler M, Bolker BM, Walker SC. Fitting linear mixed-effects models using lme4. J Stat Softw. 2015;67(1). Google Scholar
- Ritz C, Streibig JC. Dose response curves and other nonlinear curves in Weed Science and Ecotoxicology with the add-on package drc in R. Rvignette. 2012;1-51. Google Scholar
- Mulla MS, Darwazeh HA, Norland RL. Insect growth regulators: evaluation procedures and activity against mosquitoes. J Econ Entomol. 1974;67(3):329-332. PubMed | Google Scholar
- Farnesi LC, Brito JM, Linss JG, Pelajo-Machado M, Valle D, Rezende GL. Physiological and morphological aspects of Aedes aegypti developing larvae: Effects of the chitin synthesis inhibitor novaluron. PLoS One. 2012;7(1):e30363. PubMed | Google Scholar
- Clemens A. The biology of mosquitoes. 2011. Google Scholar
- Kaindoa EW, Matowo NS, Ngowo HS, Mkandawile G, Mmbando A, Finda M et al. Interventions that effectively target Anopheles funestus mosquitoes could significantly improve control of persistent malaria transmission in south-eastern Tanzania. PLoS One. 2017 May 18;12(5):e0177807. PubMed | Google Scholar
- Lwetoijera DW, Kiware SS, Mageni ZD, Dongus S, Harris C, Devine GJ et al. A need for better housing to further reduce indoor malaria transmission in areas with high bed net coverage. Parasites and Vectors. 2013 Mar 7;6:57. PubMed | Google Scholar
- Yunta C, Grisales N, Nász S, Hemmings K, Pignatelli P, Voice M et al. Pyriproxyfen is metabolized by P450s associated with pyrethroid resistance in An. gambiae. Insect Biochem Mol Biol. 2016 Nov;78:50-57. PubMed | Google Scholar
- Tusting LS, Thwing J, Sinclair D, Fillinger U, Gimnig J, Bonner KE et al. Mosquito larval source management for controlling malaria. Cochrane Database Syst Rev. 2013 Aug 29;2013(8):CD008923. PubMed | Google Scholar
- WHO. Novaluron in drinking-water: Use for vector control in drinking-water sources and containers-background document for development of WHO guidelines. WHO/SDE/WSH/07.01/11. Geneva. 2007.
Search
This article authors
On Pubmed
On Google Scholar
Citation [Download]
Navigate this article
Similar articles in
Key words
Tables and figures




Article metrics
Recently from the PAMJ








Authors´ services
